On this page a number of characteristic stages of the embryonic development in fixed
A. Fertilized egg:
0 - 1 hour. One cell stage, shortly after fertilization. The sperm penetrates the animal hemisphere mostly under an angle of about 40° with respect to the apical pole of the oocyte (not visible here). After penetration of the sperm, a grey crescent is formed at the opposite side of the point of sperm entry due to the rotation of the cortical cytoplasm. The position of the grey crescent determines the orientation of the bilateral symmetry in the egg and the developing embryo. (Not visible in Xenopus) |
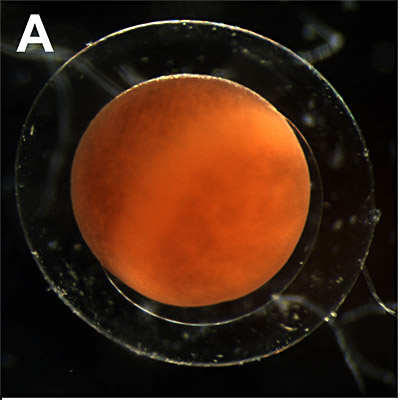 |
B. Two cells stage:
1 hour. The first cleavage occurs in a vertical plane and the cleavage furrow begins at the animal pole and slowly extends down into the vegetal region. The first cleavage plane corresponds to the plane of bilateral symmetry in the developing embryo. |
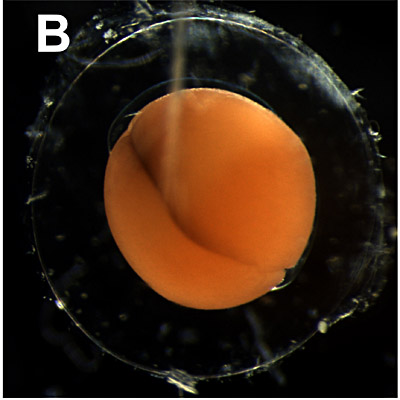 |
C Four cells stage:
2 hours. The second vertical cleavage is completed at right angles to the first one. The four formed blastomeres can differ in largeness and color. The cleavage is not always complete due to the higher concentration of yolk at the vegetal pole. | 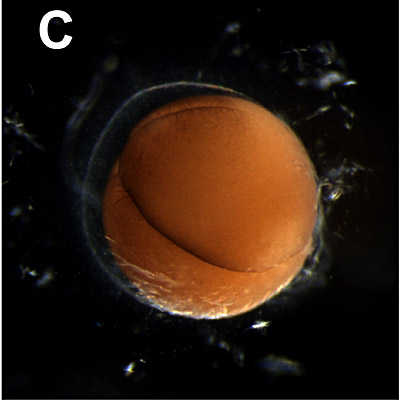 |
D. Sixteen cells stage:
2 - 3 hours. Earlier, at the eight cell stage (not shown), the third cleavage occurred horizontally and just above the equatorial plane, resulting in a difference in largeness of the blastomeres: micromeres at the animal pole and macromeres at the vegetal pole. Now, at the sixteen cell stage, the cleavage furrows are completed and the macromeres are fully separated. The dorsal micromeres are mostly smaller and less pigmented than the ventral macromeres. | 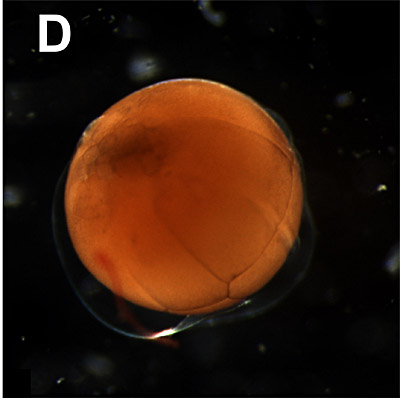 |
E. Early gastrulation:
7 - 10 hours. The gastrulation process starts after the blastula stage (not shown) in which the micromeres are sometimes difficult distinguishable as individual cells (see further in figure F). | 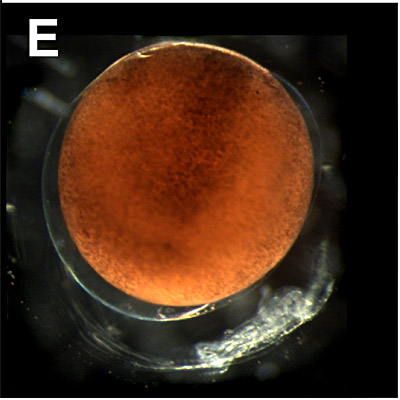 |
F. Early gastrulation (continued):
10 hours. First, the blastopore becomes visible due to so-called morphogenetic movements (migration of blastomeres). The cells at the surface of the dorsal side move inside (invaginate) at the dorsal blastopore lip. | 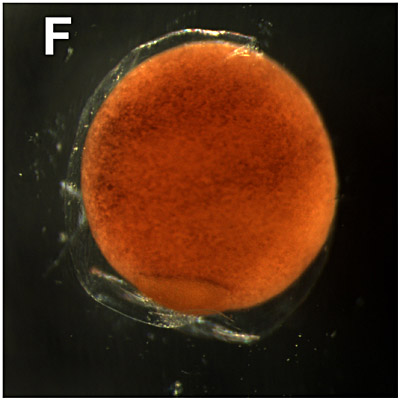 |
G. Late gastrulation/yolk plug stage:
12 hours. The circular blastopore is formed by the process of involution and epiboly whereby the dorsal, the ventral and the two lateral blastopore lips grow to each other. At the end of the gastrulation process, this opening is completely filled with a dense mass of yolk rich cells: the yolk plug. | 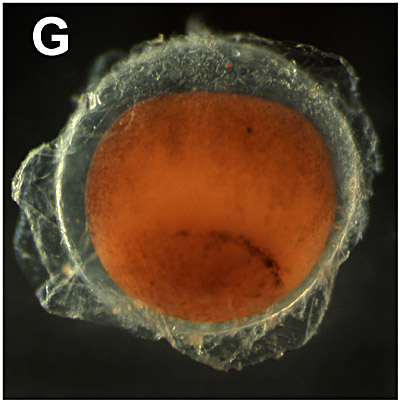 |
H. Neural groove/ Neural tube:
Between 15 and 24 hours. The neurulation process induces the formation of the nervous system. The ectoderm at the dorsal side of the gastrula thickens about 14 hours after fertilization and forms the neural plate as a presumptive structure of the nervous system. One hour later the cells of this neural plate moves inwards in the median region of the plate to form the neural groove. The neural walls develop at each side of the groove and grow to each other to form the neural tube 24 hours after fertilization. In this figure, the transition of neural groove to neural tube is shown. | 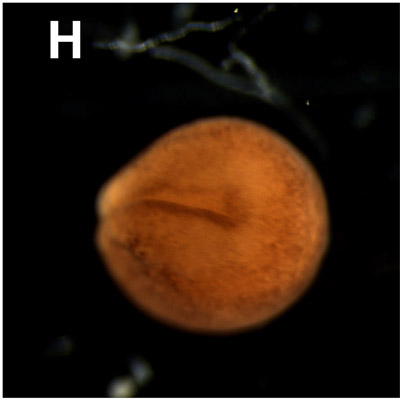 |
I. Neural tube:
After 24 hours. The anterior-posterior body axis is appearing now due to the extension of the embryo. This axis becomes clearly visible during further elongation growth of the embryo and the presumptive forebrain, eye vesicles and gill plates are developing. | 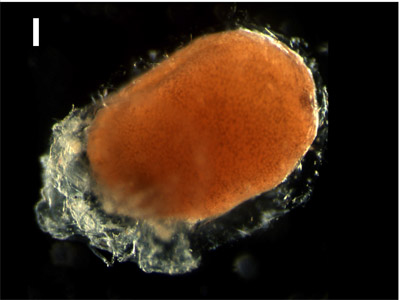 |
J. Tail bud:
1 day and 7 hours. The rudimentary eyes are clearly distinguishable. Furthermore, the place of the stomodeum (presumptive mouth) and the first external shape of the gills and tail are visible. | 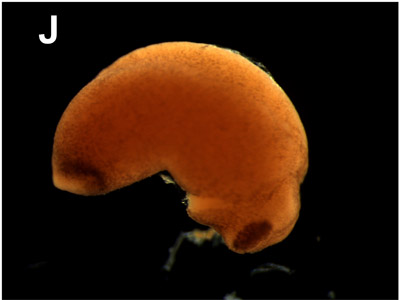 |
K. Developing larva:
The tail bud stage develops to a real larva. In this picture, we look to the dorsal side. The head, the body and the tail region are clearly visible. This larva develops further to the premetamorphose stage which will be characterized by: a broad head, two palps at both sides of the mouth, fore and back legs and a long tail laterally flattened and with a web at the dorsal and ventral side. Changes in bone, internal organs and skin appear during metamorphosis. | 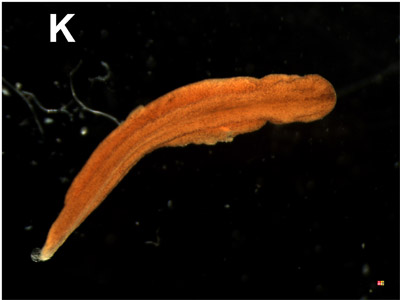 |